Main Page
|
|
|
| The Computational Memory Lab uses mathematical modeling and computational techniques to study human memory. We apply these quantitative methods both to data from laboratory studies of human memory and from electrophysiological studies involving direct human brain recordings in neurosurgical patients. Our research is focused on neurocomputational mechanisms of human episodic and spatial memory. Episodic memory refers to memory for events that are embedded in a temporal context. This includes both memory for significant life events and memory for common daily activities. In the laboratory, episodic memory is investigated by presenting lists of items (frequently words) for study, and then asking participants to recall the words. By analyzing the dynamics of the recall process one can quantify the way in which people transition from one recalled word to the next (see Fig. 1). Furthermore, by studying the electrophysiology of the brain while engaged in memory tasks, we can find, for example, regions that show increased or decreased activity when a word is successfully encoded (i.e., later recalled) versus when it is not successfully encoded, known as the subsequent memory effect (see Fig. 3). Two of our ongoing, large-scale data collection projects are the Penn Electrophysiology of Encoding and Retrieval Study (PEERS), a multi-session experiment with young and older adults combining free recall and scalp EEG (a book of these results can be found here); and an effort to collect electrophysiological data on patients with intractable epilepsy (undergoing monitoring with intracranial electrodes at partnering local hospitals) while they participate in a variety of memory and decision-making tasks. |
|

Computational models of human memory
To explain the processes underlying encoding, organization and retrieval of episodic memories, Kahana and colleagues (notably Marc Howard, Sean Polyn, Per Sederberg, and Lynn Lohnas) have developed a class of retrieved-context models. These models assume that the input to the memory system itself produces contextual drift, and that the current state of context is used to retrieve items from memory. The temporal context model (TCM; Howard and Kahana, 2002) was introduced to explain recency and contiguity effects in free recall. Specifically, recency effects appear because the context at the time of the memory test is most similar to the context associated with recent items. When an item is retrieved at test, it reinstates the context active when that item was studied. Because this context overlaps with the encoding context of the items' neighbors, a contiguity effect results. Consistent with experimental data, TCM and its variants also predict that recency and contiguity effects are approximately time-scale invariant (Sederberg, Howard, and Kahana, 2008). Polyn, Norman, and Kahana (2009) developed the Context Maintenance and Retrieval model (CMR), which is a generalized version of TCM that accounts for the influence of non-temporal associations (e.g., semantic knowledge) on recall dynamics. MATLAB scripts to run the CMR model can be downloaded here.
Despite the vast stores of memories we accumulate over a lifetime of experience, the human memory system is often able to target just the right information, seemingly effortlessly. How does the memory system accomplish this feat? Most previous models made the simplifying assumption that memory search is automatically restricted to a target list, largely bypassing the need to target the right information. Lohnas, Polyn, and Kahana (2015) developed CMR2 to address this issue. In CMR2, memory accumulates across multiple experimental lists, and temporal context is used both to focus retrieval on a target list, and to censor retrieved information when its match to the current context indicates that it was learned in a non target list. The model simultaneously accounts for a wide range of intralist and internist phenomena, including the pattern of prior-list intrusions observed in free recall, build-up of and release from proactive interference, and the ability to selectively target retrieval of items on specific prior lists (Jang & Huber, 2008; Shiffrin, 1970). Healey and Kahana (2015) used CMR2 to better understand why memory tends to get worse as we age. By fitting CMR2 to the performance of individual younger and older adults, they identified deficits in four critical processes: sustaining attention across a study episode, generating retrieval cues, resolving competition, and screening for inaccurate memories (intrusions). MATLAB scripts to run the CMR2 model can be downloaded here.
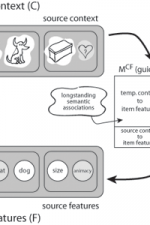
 click to enlarge Fig. 2: The context-maintenance and retrieval model. A. Schematic of CMR. When an item is studied its feature representation (fi) is activated on F. The feature representation contains both item and source features (e.g., size / animacy judgment). An associative weight matrix (MFC) allows each item to update a context representation that contains the item and source features of recently studied items. During study, the features of each item are associated with coactive context elements. During recall, the context representation reactivates (through MCF) the features of recently studied items. B. Hypothesized interactions between prefrontal cortex, temporal cortex, and medial temporal lobe during memory encoding and retrieval predicted by CMR. C. IRTs as a function of output position and total number of recalled items (4, 5, 6 or 7). D. Serial position curves for list lengths (LL) of 20, 30 and 40 items. |

Neural oscillatory correlates of episodic memory
We investigate the neurophysiology of episodic memory with electrocorticographic (ECoG) and single neuron recordings from neurosurgical patients who have had electrodes surgically implanted on the cortical surface of the brain or in the medial temporal lobes (including hippocampus) as part of the clinical process of localizing seizure foci. One focus of this research is to determine the oscillatory correlates of successful episodic memory formation and retrieval. Analyses of such recordings have shown that 65 - 95 Hz (gamma) brain oscillations increase while participants are studying words that they will successfully, as opposed to unsuccessfully, recall (Sederberg et al., 2003; Sederberg et al., 2006; Burke et al., 2014; Long et al., 2014; for a video, click here ). Gamma activity in hippocampus and neocortex likewise increases prior to successful recall (Sederberg et al., 2007; Lega et al., 2011; Burke et al., 2014; for a video, click here ). The movie below illustrates these findings.
|
Fig. 3: Free Recall Paradigm By using a free recall task in which participants study a list of words, we can measure episodic memory formation by comparing the spectral correlates associated with encoding items that are later recalled (red) or forgotten (blue). We record electroencephalographic (EEG) signals from subdurally implanted electrodes in patients with medically intractable epilepsy. We can extract spectral signals (power of a given frequency) from the raw EEG voltage traces for each item and measure when and where power at particular frequencies changes. Successful memory formation is associated with increases in gamma band (65 - 95 Hz) activity in left lateral temporal lobe, medial temporal lobe, and left prefrontal cortex. The same analyses can be performed on items during recall to assess when and where memories are retrieved. Successful memory retrieval is associated with increases in gamma band activity in the left neocortex and hippocampus as well as increases in theta band (4 -8 Hz) activity in right temporal lobe. |
The ability to reinstate this contextual information during memory search has been considered a hallmark of episodic, or event-based, memory. In Manning et al., 2011, we sought to determine whether contextual reinstatement may be observed in electrical signals recorded from the human brain during episodic recall. We examined ECoG activity from 69 neurosurgical patients as they studied and recalled lists of words in a delayed free recall paradigm (Fig. 4A), and computed similarity between the ECoG patterns recorded just prior to each recall with those recorded after the patient had studied each word. We found that, upon recalling a studied word, the recorded patterns of brain activity were not only similar to the patterns observed when the word was studied, but were also similar to the patterns observed during study of neighboring list words, with similarity decreasing reliably with positional distance (Fig. 4C), just as predicted by context reinstatement models of free recall. The degree to which individual patients exhibited this neural signature of contextual reinstatement was correlated with the contiguity effect as seen in Fig. 4D. In this way, the study provides neural evidence for contextual reinstatement in humans.
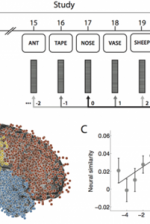
 Fig. 4: Neural evidence for contextual reinstatement in humans. A. After studying a list of 20 words and performing a brief distraction task, a participant recalls as many words as he can remember, in any order. ECoG activity is recorded during each study and recall event. The similarity between the recorded patterns is computed as a function of lag. B. Each dot marks the location of a single electrode [temporal lobe (1,815 electrodes), frontal lobe (1,737 electrodes), parietal lobe (512 electrodes), and occipital lobe (138 electrodes)]. C. Similarity between the activity recorded during recall of a word from serial position i and study of a word from serial position i + lag. (Black dot denotes study and recall of the same word, i.e., lag = 0.) D. Participants exhibiting stronger neural signatures of context reinstatement also exhibited more pronounced contiguity effects (as measured by a percentile-based temporal clustering score). (Figure after Manning et al., 2011.) |
Human spatial memory and cognition

Our lab is also interested in the neural mechanisms underlying human spatial cognition. In this work, we use virtual reality computer games in which participants learn the locations of landmarks in virtual environments. To download a sample of a YellowCab session, click here. Using this approach, we have documented the existence and character of the 4-8 Hz theta rhythm in the human brain as participants learned to navigate through complex virtual environments (Kahana et al., 1999; Caplan et al., 2001; Caplan et al., 2003; Ekstrom et al., 2005; Jacobs et al., 2010a). Recording individual neurons during virtual navigation, we have discovered "place cells" in the human brain. These cells, which are found primarily in the human hippocampus, become active when a given spatial location is being traversed Ekstrom et al. (2003). We also identified several other cellular responses during navigation: cells that become active in response to viewing a salient landmark (from any location), cells that become active when searching for a particular goal location (irrespective of location or view), and cells that respond when traveling in a given direction (bearing/heading).
Jacobs et al. (2010b) examined recordings of single-neuron activity from neurosurgical patients playing a virtual-navigation video game. In addition to place cells, which encode the current virtual location, we describe a unique cell type, entorhinal cortex (EC) path cells, the activity of which indicates whether the patient is taking a clockwise or counterclockwise path around the virtual square road. We find that many EC path cells exhibit this directional activity throughout the environment, in contrast to hippocampal neurons, which primarily encode information about specific locations. More broadly, these findings support the hypothesis that EC encodes general properties of the current context (e.g., location or direction) that are used by hippocampus to build unique representations reflecting combinations of these properties.
Grid cells in the entorhinal cortex appear to represent spatial location via a triangular coordinate system. Such cells, which have been identified in rats, bats and monkeys, are believed to support a wide range of spatial behaviors. Recording neuronal activity from neurosurgical patients performing a virtual-navigation task, Jacobs et al. (2013) identified cells exhibiting grid-like spiking patterns in the human brain, suggesting that humans and simpler animals rely on homologous spatial-coding schemes.
Theories of episodic memory suggest that memory encoding and retrieval are facilitated by spatiotemporal context: a continually updated representation of location in space and time. Recent work in the lab sought to examine the role of spatial context in human episodic memory retrieval through a hybrid spatial and episodic memory task. Miller et al. (2013) identified place cells as neurosurgical patients performed a delivery task which required them to deliver items to different stores in a virtual town. Following this navigation task, patients were asked to freely recall the items they had delivered. Place cell activation patterns during the navigational task were then compared to the subsequent activation patterns that occurred during the free recall task. Miller et al. found that neural activity during the retrieval of each delivered item was similar to the neural activity associated with the location where that item was encoded. These findings demonstrate context-specific reinstatement of place-responsive cell activity at the time of recall, supporting theories that implicate contextual reinstatement as the basis for memory retrieval.
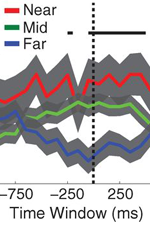
 Fig. 8: Spatial context reinstatement. (a) Time courses of neural similarity between ensemble place-cell activity during navigation and during item recall for near, middle, and far spatial distance bins. Shaded regions indicate SEM across recalled items, the horizontal bars indicate statistically significant time points and the vertical dotted line indicates the onset of vocalization. (b) Average neural similarity for near, middle and far distance bins for the time period of -300 to 700ms relative to recall onset. Error bars indicate SEM across recalled items. (c) Neural similarity for near and far spatial distance bins for each participants (colored lines) and the participant average (black line) during -300 to 700ms relative to recall onset. Error bars indicate SEM across participants. |