Sandbox
|
|
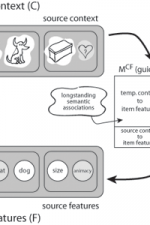
Computational models of human memoryTo explain the processes underlying encoding, organization and retrieval of episodic memories, Kahana and colleagues (notably Marc Howard, Sean Polyn, Per Sederberg, and Lynn Lohnas) have developed a class of retrieved-context models. These models assume that the input to the memory system itself produces contextual drift, and that the current state of context is used to retrieve items from memory. The temporal context model (TCM; Howard and Kahana, 2002) was introduced to explain recency and contiguity effects in free recall. Specifically, recency effects appear because the context at the time of the memory test is most similar to the context associated with recent items. When an item is retrieved at test, it reinstates the context active when that item was studied. Because this context overlaps with the encoding context of the items' neighbors, a contiguity effect results. Consistent with experimental data, TCM and its variants also predict that recency and contiguity effects are approximately time-scale invariant (Sederberg, Howard, and Kahana, 2008). Polyn, Norman, and Kahana (2009) developed the Context Maintenance and Retrieval model (CMR), which is a generalized version of TCM that accounts for the influence of non-temporal associations (e.g., semantic knowledge) on recall dynamics. MATLAB scripts to run the CMR model can be downloaded here. Despite the vast stores of memories we accumulate over a lifetime of experience, the human memory system is often able to target just the right information, seemingly effortlessly. How does the memory system accomplish this feat? Most previous models made the simplifying assumption that memory search is automatically restricted to a target list, largely bypassing the need to target the right information. Lohnas, Polyn, and Kahana (2015) developed CMR2 to address this issue. In CMR2, memory accumulates across multiple experimental lists, and temporal context is used both to focus retrieval on a target list and to censor retrieved information when its match to the current context indicates that it was learned in a non-target list. The model simultaneously accounts for a wide range of intralist and interlist phenomena, including the pattern of prior-list intrusions observed in free recall, build-up of and release from proactive interference, and the ability to selectively target retrieval of items on specific prior lists (Jang & Huber, 2008; Shiffrin, 1970). Healey and Kahana (2015) used CMR2 to better understand why memory tends to get worse as we age. By fitting CMR2 to the performance of individual younger and older adults, they identified deficits in four critical processes: sustaining attention across a study episode, generating retrieval cues, resolving competition, and screening for inaccurate memories (intrusions). Healey and Kahana extended CMR2 to simulate a recognition memory task using the same mechanisms the free recall model uses to reject intrusions. Without fitting any additional parameters, the model accounts for age differences in recognition memory accuracy. Confirming a prediction of the model, free recall intrusion rates correlate positively with recognition false alarm rates. MATLAB scripts to run the CMR2 model can be downloaded here.
Neural oscillatory correlates of episodic memoryWe investigate the neurophysiology of episodic memory with electrocorticographic (ECoG) and single neuron recordings from neurosurgical patients who have had electrodes surgically implanted on the cortical surface of the brain or in the medial temporal lobes (including hippocampus) as part of the clinical process of localizing seizure foci. One focus of this research is to determine the oscillatory correlates of successful episodic memory formation and retrieval. Analyses of such recordings have shown that 65 - 95 Hz (gamma) brain oscillations increase while participants are studying words that they will successfully, as opposed to unsuccessfully, recall (Sederberg et al., 2003; Sederberg et al., 2006; Burke et al., 2014; Long et al., 2014; Long and Kahana, 2015; for a video, click here ). Gamma activity in hippocampus and neocortex likewise increases prior to successful recall (Sederberg et al., 2007; Lega et al., 2011; Burke et al., 2014; for a video, click here ). The movie below illustrates these findings.
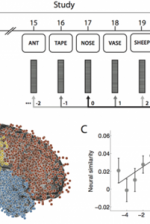
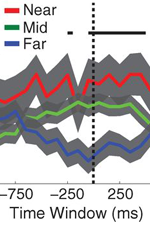
The ability to reinstate this contextual information during memory search has been considered a hallmark of episodic, or event-based, memory. In Manning et al., 2011, we sought to determine whether contextual reinstatement may be observed in electrical signals recorded from the human brain during episodic recall. We examined ECoG activity from 69 neurosurgical patients as they studied and recalled lists of words in a delayed free recall paradigm (Fig. 4A), and computed similarity between the ECoG patterns recorded just prior to each recall with those recorded after the patient had studied each word. We found that, upon recalling a studied word, the recorded patterns of brain activity were not only similar to the patterns observed when the word was studied, but were also similar to the patterns observed during study of neighboring list words, with similarity decreasing reliably with positional distance (Fig. 4C), just as predicted by context reinstatement models of free recall. The degree to which individual patients exhibited this neural signature of contextual reinstatement was correlated with the contiguity effect as seen in Fig. 4D. In this way, the study provides neural evidence for contextual reinstatement in humans.
Human spatial memory and cognitionOur lab is also interested in the neural mechanisms underlying human spatial cognition. In this work, we use virtual reality computer games in which participants learn the locations of landmarks in virtual environments. To download a sample of a YellowCab session, click here. Using this approach, we have documented the existence and character of the 4-8 Hz theta rhythm in the human brain as participants learned to navigate through complex virtual environments (Kahana et al., 1999; Caplan et al., 2001; Caplan et al., 2003; Ekstrom et al., 2005; Jacobs et al., 2010a). Recording individual neurons during virtual navigation, we have discovered "place cells" in the human brain. These cells, which are found primarily in the human hippocampus, become active when a given spatial location is being traversed (Ekstrom et al., 2003). We also identified several other cellular responses during navigation: cells that become active in response to viewing a salient landmark (from any location), cells that become active when searching for a particular goal location (irrespective of location or view), and cells that respond when traveling in a given direction (bearing/heading). Jacobs et al. (2010b) examined recordings of single-neuron activity from neurosurgical patients playing a virtual-navigation video game. In addition to place cells, which encode the current virtual location, we describe a unique cell type, entorhinal cortex (EC) path cells, the activity of which indicates whether the patient is taking a clockwise or counterclockwise path around the virtual square road. We find that many EC path cells exhibit this directional activity throughout the environment, in contrast to hippocampal neurons, which primarily encode information about specific locations. More broadly, these findings support the hypothesis that EC encodes general properties of the current context (e.g., location or direction) that are used by hippocampus to build unique representations reflecting combinations of these properties. Grid cells in the entorhinal cortex appear to represent spatial location via a triangular coordinate system. Such cells, which have been identified in rats, bats and monkeys, are believed to support a wide range of spatial behaviors. Recording neuronal activity from neurosurgical patients performing a virtual-navigation task, Jacobs et al. (2013) identified cells exhibiting grid-like spiking patterns in the human brain, suggesting that humans and simpler animals rely on homologous spatial-coding schemes. Theories of episodic memory suggest that memory encoding and retrieval are facilitated by spatiotemporal context: a continually updated representation of location in space and time. Recent work in the lab sought to examine the role of spatial context in human episodic memory retrieval through a hybrid spatial and episodic memory task. Miller et al. (2013) identified place cells as neurosurgical patients performed a delivery task which required them to deliver items to different stores in a virtual town. Following this navigation task, patients were asked to freely recall the items they had delivered. Place cell activation patterns during the navigational task were then compared to the subsequent activation patterns that occurred during the free recall task. Miller et al. found that neural activity during the retrieval of each delivered item was similar to the neural activity associated with the location where that item was encoded. These findings demonstrate context-specific reinstatement of place-responsive cell activity at the time of recall, supporting theories that implicate contextual reinstatement as the basis for memory retrieval.
Cognitive NeuromodulationThe lab is also interested in how electrical stimulation can be used to therapeutically modulate memory function. The RAM project, supported by the Defense Advanced Research Projects Agency (DARPA), is a large-scale collaboration in which we use intracranial recording and stimulation to understand and affect memory function. By aggregating data collection from intracranial patients at many clinical centers, we are able to study in detail the neural mechanisms of memory encoding and retrieval across four tasks that address many core memory processes: free recall, categorized free recall, paired associate learning and spatial navigation. During performance of these tasks, we apply targeted electrical stimulation to specific nodes in the memory network, including the hippocampus, medial temporal lobe cortices and the frontal cortex. We then use multivariate machine learning methods to analyze the effects of stimulation on brain activity, and their relationship to memory performance. Our ultimate goal is to use this knowledge to understand how electrical stimulation can be used to treat memory dysfunction. |